



Current project
…about the roles of small axons, whose size matches that of the majority cortical axons
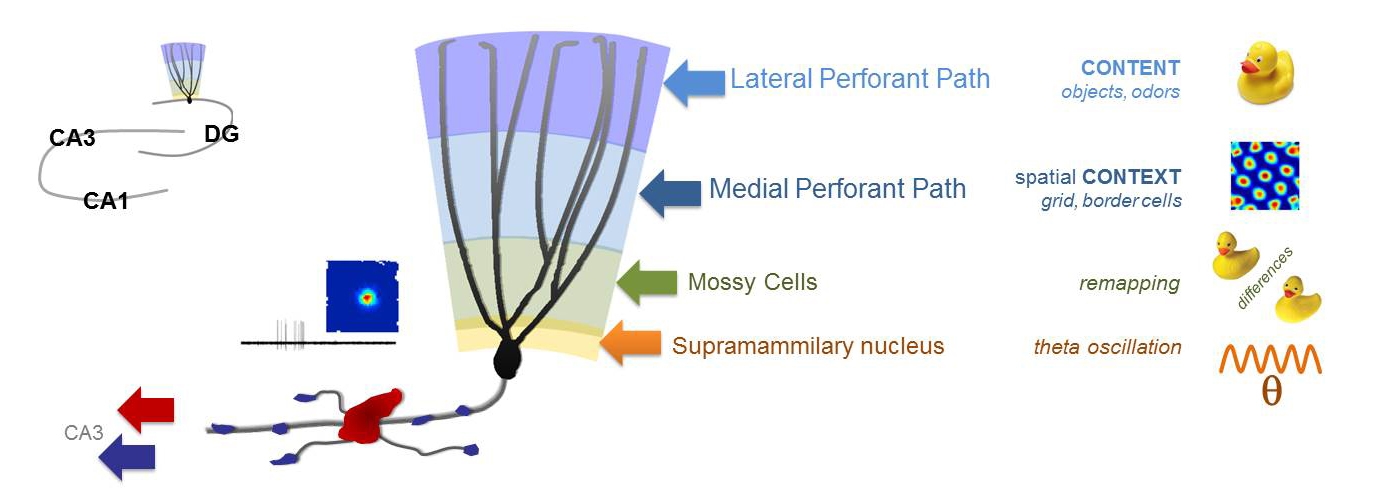
The current theories of the broader hippocampal network explain its memory functions based on the firing of individual neurons. For example, the sparse activity of dentate gyrus granule cells (DG GC) codes and relays very specific information about the local environment. However, the mechanisms that generate the information-rich GC activity are elusive, because the firings of neurons, which project to and activate GCs, are usually dense and codes different aspects of the ongoing and previous experiences. The translation of diverse inputs into highly different specific codes appears to be one of the most fundamental neuronal computation rules throughout the brain. Therefore, we must understand the principles that govern how single GCs extract information from the activity of single upstream neurons. The fact that our knowledge about the GC output is much more advanced than the excitatory drives to the GCs, are in large part due to the mechanistic insights that were gained by the direct recording techniques from the unusually large axon terminals of the GCs. Therefore, to understand the excitation of GCs at a similar, we directly record from small axon terminals that excite GCs by further improving the sophisticated axonal recordings.
Using direct patch clamp recordings and voltage-sensitive dye/protein imaging of single small axon terminals we investigate the functional contributions of the axonal excitability properties of the major glutamatergic inputs to the DG region. We also investigate the output that they generate in DG granule cells, and the mechanisms of relaying this along their specialized axon, the so-called mossy fibers. Mossy fibers form both typical sized axon terminals and also some unusually large terminals that are funcitonally distinct as their innervate distinct postsynaptic neurons. We investigate how the different biophysical properties of small and large terminals affect action potential propagation.
Direct electrophysiology methods give several advantages. With whole bouton recordings, we can measure fundamental signaling properties, including the exact spike shape, which is important for how much calcium enters and how reliable is the subsequent synaptic release. Or we can also observe the plasticity of spike shapes during physiological activities. We identify the recorded axons with posthoc morphological analysis and immuno-labelling. By pulling outside out patches from small axons, we can understand the underlying ion-channel mechanisms of their firing. Using additional methods we make sure that the recorded signals are correct and not contaminated by the influence of the recording pipettes, including voltage sensitive dye imaging and simulations, where the axonal structure, ionic conductances and also the recording instruments are accurately implemented.
{kind=link}
Furthermore, combination of in vitro slice recordings with imaging of GEVI signals (genetically encoded voltage indicators) allows the measurement of the unitary synaptic responses, similarly to classical paired recording, and map synaptic connectivity in a large neuronal network.