




Az agyi területek működési módjainak összehangolása II: a kéreg alatti modulációs központok szerepe
A GNW kísérleti megalapozását már áttekintettük, és nagyjából értjük a tudattartalmak dinamikáját. A kérdés azonban továbbra is nyitott: Hogyan képes a homloklebeny más agyterületeket — elsősorban az agykérget — összehangolt működésű hálózatokba szervezni?
Az anatómiai kapcsolatok alapján legalább három lehetséges „meghívó” útvonal rajzolódik ki:
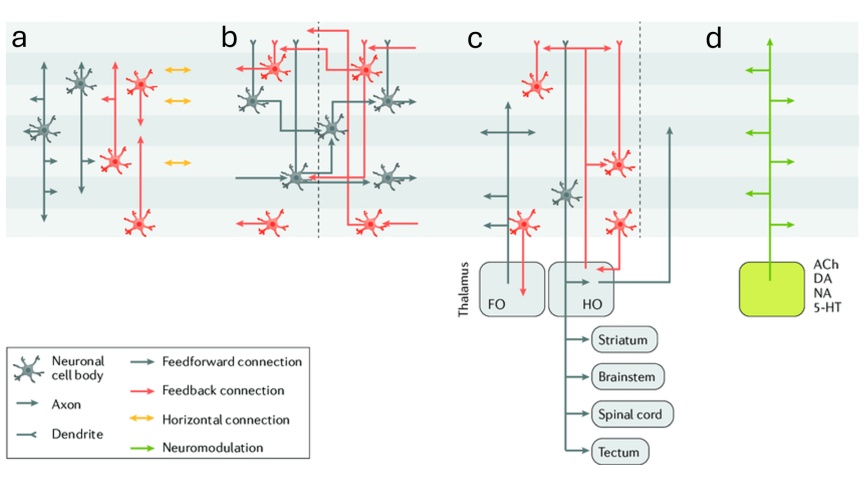
1) Kéreg–kéreg (cortico-corticalis) kapcsolatok
Az agykérgi területek között kiterjedt, hosszú távú serkentő pályák futnak, melyek nagy mennyiségű információt továbbítanak. Ezek két alapvető irányt követnek:
• Feedforward kapcsolatok: főként a II–III. rétegből indulnak, és a célterületek IV. rétegében végződnek. Ezek közvetítik az alacsonyabb szintről magasabb szintre haladó információt.
• Feedback kapcsolatok: jellemzően az V–VI. rétegből indulnak, és a célterületek I. és VI. rétegében végződnek. Ezek top-down hatásokat közvetítenek.
Ez a kettős rendszer teszi lehetővé, hogy a magasabb rendű területek ne csak „olvassák”, hanem aktívan alakítsák is az alacsonyabb szintű feldolgozást — például a releváns modell kiválasztásán keresztül.
2) Kéreg–bazális ganglion–talamusz hurkok
A kéregből induló jelek a striátumon és a bazális ganglionokon keresztül jutnak vissza a kéregbe a talamusz közvetítésével. Itt két eltérő működésű thalamikus rendszer különíthető el:
• Specifikus (core, FO) projekciók, amelyek a IV. rétegbe érkeznek, és klasszikus feedforward jellegűek.
• Diffúz (matrix, HO) projekciók, amelyek széles körben, elsősorban az I. rétegbe vetítenek, és inkább globális állapotszabályozó szerepet töltenek be.
Ez a rendszer nem csupán információt továbbít, hanem szelekciót is végez: eldönti, hogy mely kérgi aktivitásmintázatok erősödjenek meg és kerüljenek vissza a kéregbe.
3) Kéreg alatti moduláló rendszerek
A harmadik útvonal minőségileg eltér az előző kettőtől. Az agytörzsi és bazális előagyi modulátor rendszerek a gyors szinaptikus transzmisszióban szerepet játszó serkentő glutamát és gátló GABA mellett egyedi transzmittereket használnak, melyek felszabadulása hangolja a többi agyterület sejtjeinek aktivitását, így nem elsősorban konkrét információt, hanem állapotot közvetítenek. A mediális szeptum és bazális előagy acetil-kolint, a raphe magvak szerotonint, a locus coeruleus noradrenalint, a substancia nigra és ventral tegmentális terület dopamint, a tuberomamilláris mag pedig hisztamint használ, hogy:
• szabályozzák a neuronok ingerlékenységét,
• befolyásolják a jel–zaj viszonyt,
• módosítják a szinaptikus hatékonyságot,
• és kulcsszerepet játszanak a tanulásban.
Fontos hangsúlyozni, hogy nem pusztán lassú, diffúz hatásokról van szó. A modulátor rendszerek tonikus és fázisos komponensekkel is működnek: képesek gyors, időben pontos jelzésekre (például dopamin alapú predikciós hibák), miközben hosszabb időskálán is hangolják az agyi állapotokat. Károsodásuk súlyos tudatzavarhoz vezethet (csökkent, beszűkölt tudatállapot, tudatvesztés), ami arra utal, hogy ezek a rendszerek szükséges feltételei a tudatos működésnek — még ha a konkrét tartalmat nem is ők hozzák létre.
Ezek a moduláló központok (MK a továbbiakban) az evolúció során korán kialakult ősi agyterületeken találhatók, és a kezdetektől részt vettek a szervezet belső állapotai által meghatározott igények alapján az agyműködés hangolásában. Ahogy kialakult és bonyolódott az agykéreg, az MK-k kérgi vetítéseik útján rajta tartották a kezüket, hogy működését a szervezet és a környezet igényeinek megfelelően hangolják, ne hagyják a fellegekben szárnyalni, mert abba bele lehet halni. Így meghatározóvá váltak a külső környezet és a belső állapotok változását követő állapotátmentek hangolásában, a figyelemben, az éberségben, az alvás-ébrenléti ciklus szabályozásába és a motivációban.
De miért és hogyan meghatározók az MK-k a tudathoz szükséges dinamikus agykérgi hálózati kölcsönhatások kialakításában?
Korábban amellett érveltem, hogy egy idegi hálózat nem képes egyszerre minden feladatot optimálisan ellátni. Egy agykérgi területnek időben váltogatnia kell különböző működési módokat — például információ fogadása, feldolgozása és továbbítása között. Amikor több terület működik együtt, ezeknek a váltásoknak időben összehangoltnak kell lenniük: a küldő területnek „kimeneti” állapotba kell kerülnie, a fogadónak pedig ezzel szinkronban „bemeneti” állapotba. Többek között kutatócsoportunk munkái mutatták ki (lásd a Tengericsikó meséjét), hogy a moduláló ingerületátvivő anyagokat felszabadító MK-k fontosak az agyi feldolgozási módok, és ezért az EEG állapotok közötti átkapcsolásban. Ha az MK-kból érkező jelek nem egyszerűen aktiválnak, hanem időben és térben finoman strukturált mintázatokban hangolják a kérgi területek működési módját, összerendezhetik a kéregterületeket közötti információ áramlását. Ez a gyorsan változó hangolás nyilvánul meg az EEGben összehangolt gamma, béta, theta oszcillációként. Ehhez persze az kell, hogy a moduláló bemenet az egyes célterületeket ne egyszerre kapcsolják ugyanolyan állapotba, hanem egy finom, változó mintázatban. 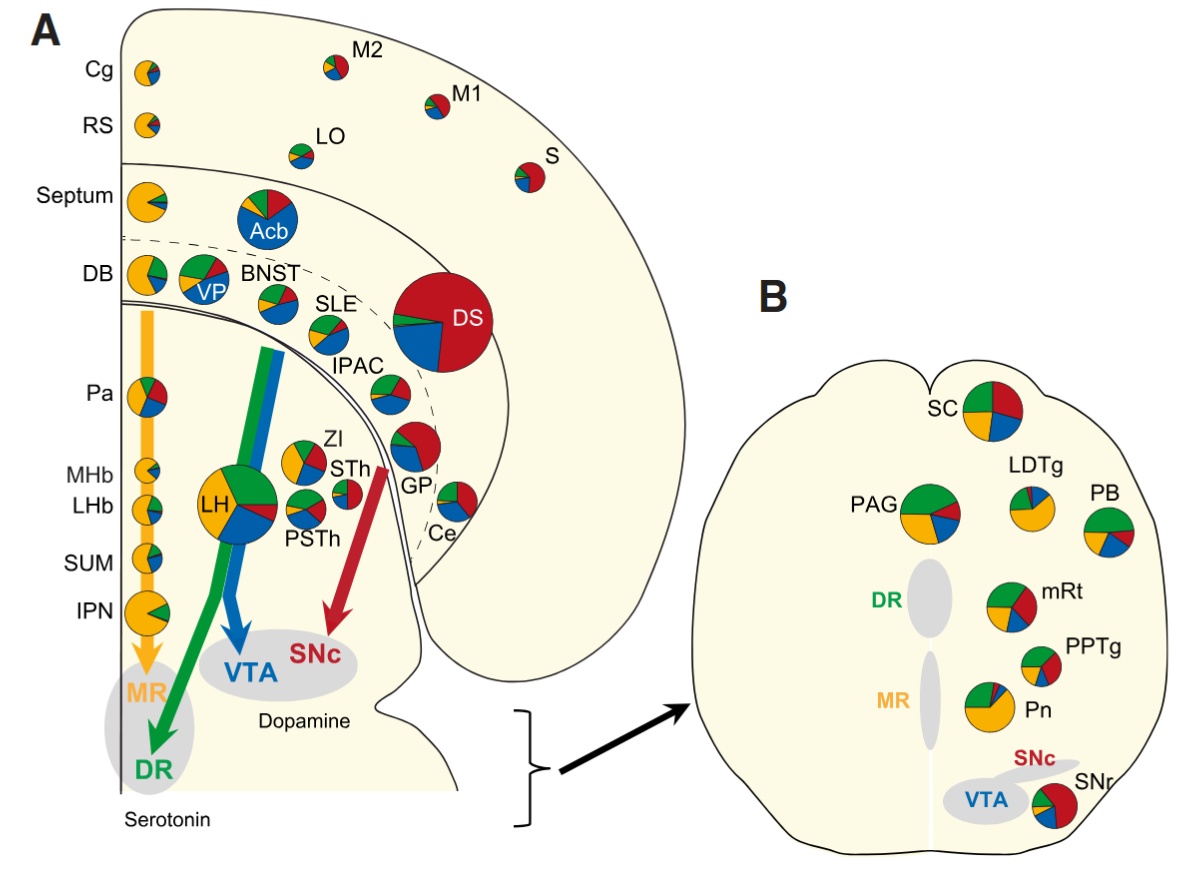 Ezt a lehetőséget támogatják a legújabb eredmények. Korábban az volt az uralkodó nézet, hogy ezek a rendszerek szétszórtan, nem specifikusan árasztják el az agyat. Az újabb eredmények ezt a képet jelentősen finomították. Ma már tudjuk, hogy: egy-egy modulátor magon belül több sejttípus létezik, ezek eltérő bemeneteket kapnak és gyakran jól körülhatárolt célterületekre vetítenek. Például a dopaminerg rendszerben a VTA egyedi neuronjai specifikus kérgi és striátális célpontokat érnek el. Hasonló szerveződés figyelhető meg a kolinerg és szerotonerg rendszerekben is. Ez a strukturáltság teszi lehetővé, hogy a különböző kérgi területek ne egyszerre, hanem finoman eltolva, mintázatban váltsanak működési módot.
Ezt a lehetőséget támogatják a legújabb eredmények. Korábban az volt az uralkodó nézet, hogy ezek a rendszerek szétszórtan, nem specifikusan árasztják el az agyat. Az újabb eredmények ezt a képet jelentősen finomították. Ma már tudjuk, hogy: egy-egy modulátor magon belül több sejttípus létezik, ezek eltérő bemeneteket kapnak és gyakran jól körülhatárolt célterületekre vetítenek. Például a dopaminerg rendszerben a VTA egyedi neuronjai specifikus kérgi és striátális célpontokat érnek el. Hasonló szerveződés figyelhető meg a kolinerg és szerotonerg rendszerekben is. Ez a strukturáltság teszi lehetővé, hogy a különböző kérgi területek ne egyszerre, hanem finoman eltolva, mintázatban váltsanak működési módot.
Viszont amit idáig leírtunk, az egy kéreg-alatti -> agykéreg információ áramlási irány. Mindaddig, amíg a kéreg alatti területeknek csak a külvilágból és a belső testi dolgokról érkező ingerekre kell reagálni, ezzel az egyirányúsággal nem is lenne gond. De a fejlettebb agyú állatokban, ahol a cselekvés nem modell-független, hanem egy összetett belső modell irányítja (emlősök, bizonyára a madarak és valószínűleg néhány puhatestű és rák) az agykéreg (vagy az annak megfelelő terület) működését és területeinek összehangoltságát absztraktabb, csak az agykéregben (vagy azzal ekvivalens struktúrákban) kiértékelhető információk is befolyásolják.
De hogyan kerül le ez a kifinomult agykérgi információ a kéreg alatti területekre?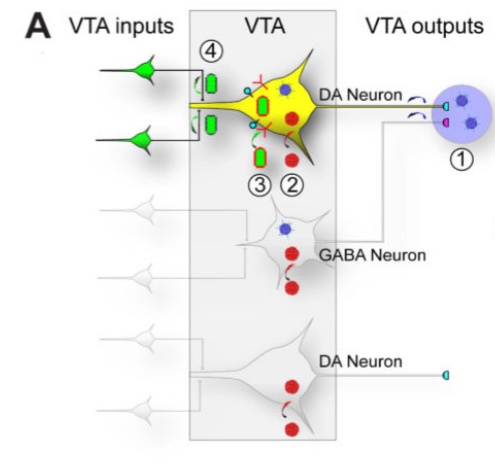 Ezt csak mostanában kezdték el megkérdezni a kutatók. A téma napjaink egyik forrongó kutatási irányává vált, melyet az tett lehetővé, hogy transzgénikus és vírus jelölési technikákat használva, lehetséges egyedi idegsejtek bemeneteinek és kimeneteinek feltérképezése. Meg tudjuk nézni azt, hogy pl. egy-egy VTA dopamin sejt, vagy a raphe magvak egyes szerotonint tartalmazó sejtjei honnan kapnak bemeneteket és hova küldik a dopamint és a szerotonint. Az agykéreg mellett, nagy erősségű (7T) fMRI -t használva elkezdték a kéreg alatti, középagyi és agytörzsi aktivitást is megvizsgálni. Itt is találtak bonyolult mintázatokat, melyek összefüggenek az agyi funkcionális hálózatok aktiválódásával.
Ezt csak mostanában kezdték el megkérdezni a kutatók. A téma napjaink egyik forrongó kutatási irányává vált, melyet az tett lehetővé, hogy transzgénikus és vírus jelölési technikákat használva, lehetséges egyedi idegsejtek bemeneteinek és kimeneteinek feltérképezése. Meg tudjuk nézni azt, hogy pl. egy-egy VTA dopamin sejt, vagy a raphe magvak egyes szerotonint tartalmazó sejtjei honnan kapnak bemeneteket és hova küldik a dopamint és a szerotonint. Az agykéreg mellett, nagy erősségű (7T) fMRI -t használva elkezdték a kéreg alatti, középagyi és agytörzsi aktivitást is megvizsgálni. Itt is találtak bonyolult mintázatokat, melyek összefüggenek az agyi funkcionális hálózatok aktiválódásával.
Az eredmények szerint, a magasan absztrakt mintázatokat feldolgozó homloklebeny különböző területeiről finoman mintázott vetítéseken keresztül nagy mennyiségű információ vetül a dopaminerg (VTA), a noradrenerg (LC), és a szerotonerg (raphe) rendszerek felé. 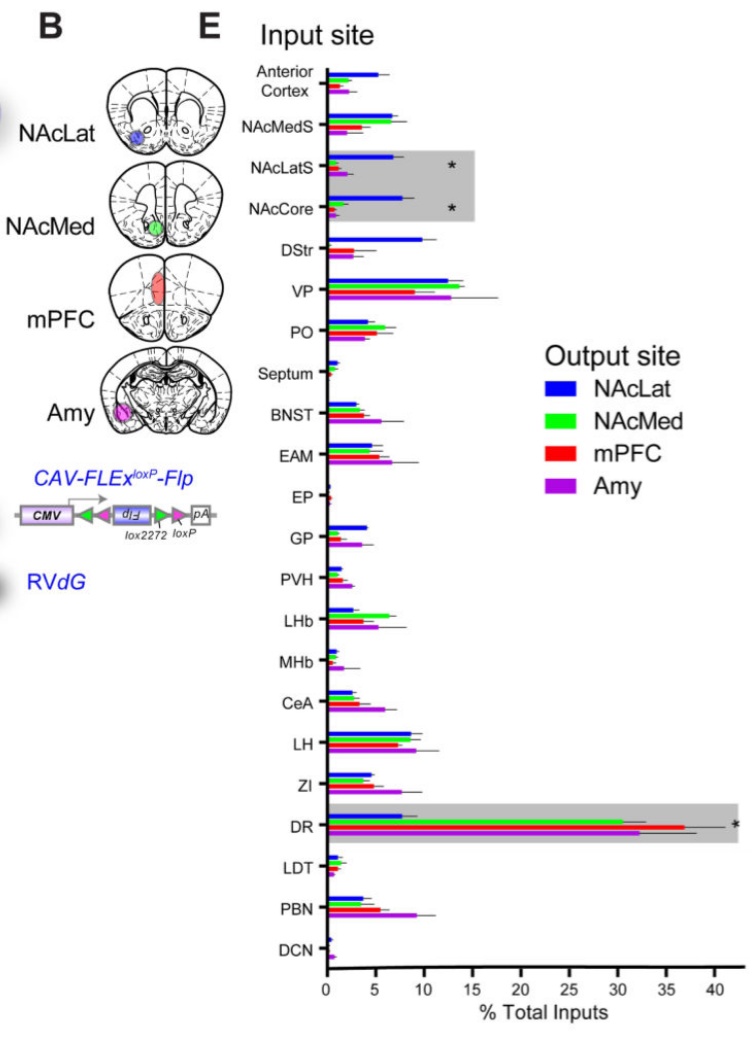 Sok MK sejtet összevetve kiderült, hogy nagy különbség van az egyedi sejtek összeköttetés rendszerében. Az, hogy mely kéregterületről kap egy sejt információt, meghatározza, hogy mely területet modulál vetítései útján. A homloklebeny tehát az MK-k, pontosabban finoman kiválasztott MK sejtek útján közvetve befolyásolni képes más agykérgi területek térbeli és időbeli kölcsönhatását, információ feldolgozó módjuk időzített átkapcsolását, azaz funkcionális hálózatok időleges kialakulását.
Sok MK sejtet összevetve kiderült, hogy nagy különbség van az egyedi sejtek összeköttetés rendszerében. Az, hogy mely kéregterületről kap egy sejt információt, meghatározza, hogy mely területet modulál vetítései útján. A homloklebeny tehát az MK-k, pontosabban finoman kiválasztott MK sejtek útján közvetve befolyásolni képes más agykérgi területek térbeli és időbeli kölcsönhatását, információ feldolgozó módjuk időzített átkapcsolását, azaz funkcionális hálózatok időleges kialakulását.
Fontos még megjegyezni, hogy a fenti három lehetséges „meghívó” útvonal nem kizárja egymást, hanem együttműködik. A reprezentációban fontos serkentő sejt mintázatok (aktiválódott modellek) az első és második útvonalon jutnak el más területekre, de ahhoz, hogy ezek a megfelelő pillanatban induljanak és érkezzenek a harmadik útvonal modulátoros hatása kapcsolja a szükséges állapotokba az aktuális funkcionális hálózatban résztvevő agyterületeket. Így jön létre a GNW-nek egy-egy pillanatnyi állapota, hálózata. Amikor egy aktuális hálózatban megszületik a megoldás, a homloklebeny annak függvényében, hogy az mi, egy újabb együttműködő hálózatot alakít ki a 3 lehetséges csatornát használva.
 Az agykérgi serkentő kapcsolatok magas információtartalmának és a moduláló központok átkapcsoló jeleinek szerepét jól lehet egy TV példájával szemléltetni. A hálózati, HDMI, és egyéb kábeleken érkezik az a rengeteg információ, hogy melyik műsort lehet megjeleníteni, de hogy be van-e kapcsolva a TV, melyik csatorna szól, és milyen hangosan, azt már a távirányítóból érkező viszonylag csekély mennyiségű információt hordozó jelek befolyásolják.
Az agykérgi serkentő kapcsolatok magas információtartalmának és a moduláló központok átkapcsoló jeleinek szerepét jól lehet egy TV példájával szemléltetni. A hálózati, HDMI, és egyéb kábeleken érkezik az a rengeteg információ, hogy melyik műsort lehet megjeleníteni, de hogy be van-e kapcsolva a TV, melyik csatorna szól, és milyen hangosan, azt már a távirányítóból érkező viszonylag csekély mennyiségű információt hordozó jelek befolyásolják.
A modern agyevolúció során tehát a homloklebeny „rátalált” az MK-kra és segítségül hívta őket az agyműködés szabályozásához. A moduláló rendszerek így már nem csak a test és a külvilág felől kapnak irányító bemeneteket, hanem a belső modellek, célok és döntési folyamatok is közvetlenül befolyásolják, hogy az MK-k mikor, hol és milyen mintázatban fejtik ki hatásukat. Egy (újabb) zárt hurok jött létre: a kéreg nemcsak használja, hanem finom felbontásban irányítja is a saját hangolását. 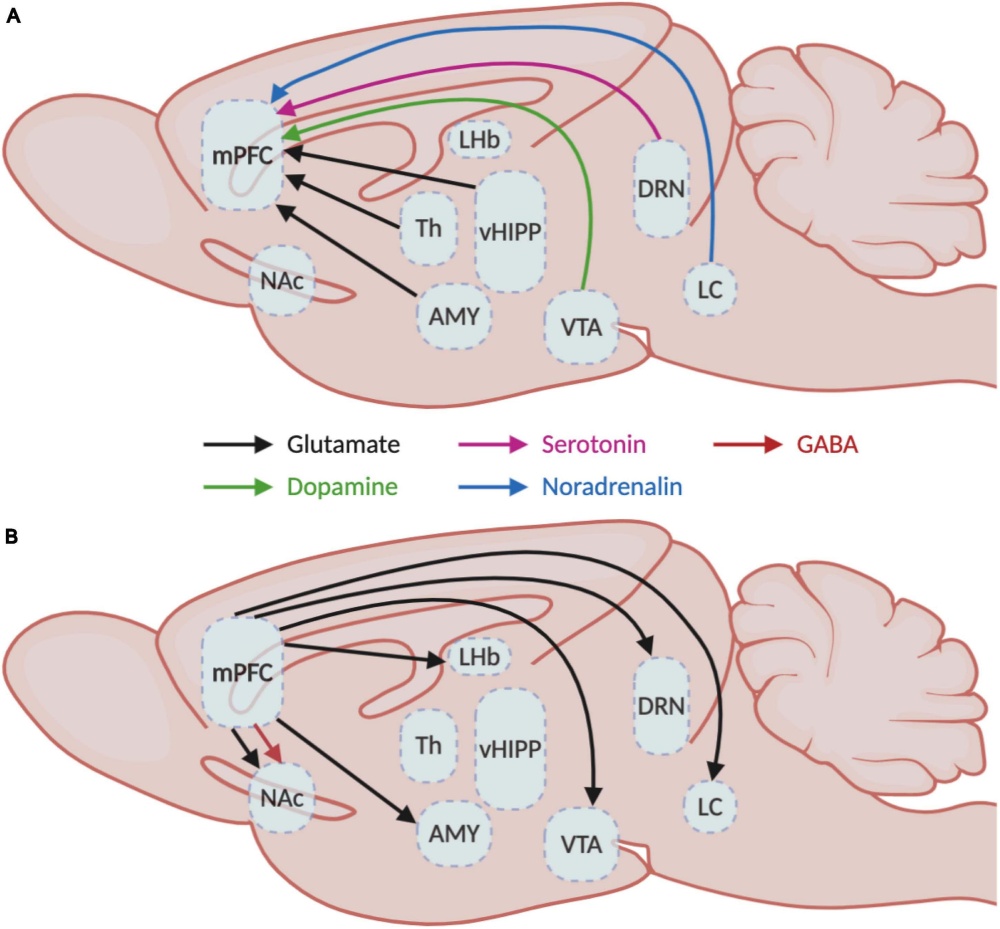 Ennek a huroknak a ciklusai felelnek meg a GNW egyes állapotainak és a hurok működése a gyorsan váltakozó tudatállapotoknak. Egy reaktív, inger vezérelt egyszerű agyból egy proaktív, motivációk és tervek által vezérelt agy lett. Úgy érzem az agykutatásban ezen a területen még sok izgalmas kérdés van, és az elkövetkező években itt várható számos nagy kátyú betömése.
Ennek a huroknak a ciklusai felelnek meg a GNW egyes állapotainak és a hurok működése a gyorsan váltakozó tudatállapotoknak. Egy reaktív, inger vezérelt egyszerű agyból egy proaktív, motivációk és tervek által vezérelt agy lett. Úgy érzem az agykutatásban ezen a területen még sok izgalmas kérdés van, és az elkövetkező években itt várható számos nagy kátyú betömése.
Ezzel végére is értünk az utolsó meredek kaptatónak mely a negyedik, az agyi meta-hálózatok dinamikájáról szóló részhez tartozott. Gratulálok mindenkinek, aki felkapaszkodott idáig. Viszont nem fejezzük még be az #agysétát, mert sok megbeszélni és finomítani valónk van. Mostantól kezdve kevesebb tény, több elmélkedés, azaz egy laposabb terep következik. Egy elméleti kitekintéssel folytatjuk, hamarosan.
Szerző: Gulyás Attila