



A bonyolult neuron modell II
Az előző bejegyzésben bemutatott új szereplők, a további ioncsatornák, a jelterjedésre két ellentétes irányban hathatnak. Amennyiben a serkentés (depolarizáció) hatására további Ca2+ és Na+ csatornák nyílnak meg, akkor ezek felerősítik a serkentés hatását. A beérkező jelek a terjedés során felerősödnek, hatékonyabban aktiválják a sejtet.
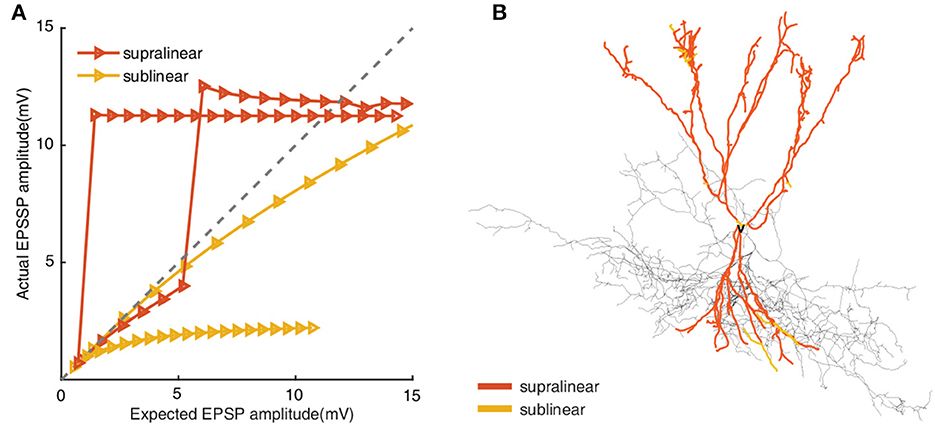 Ha egyszerre két bemenetre érkezik serkentés, hatásuk az erősítés miatt nagyobb lesz, mint a két bemenet egyszerű összege, emiatt ezt supra-lineáris összegződésnek nevezik. Olyan is lehetséges, hogy a dendrit egyik részére érkező bemenet csak akkor jut tovább, amikor a sejttesthez közelebbi dendritrészen beindulnak ezek az erősítő rendszerek. Így az egyik agyterületről származó jel csak akkor tud beterjedni, ha egy másik agyterületről megfelelő pillanatban szintén érkezik információ. Ezt az információ kapuzásának nevezik, hiszen az egyik bemenet nyitja meg a kaput a másik számára.
Ha egyszerre két bemenetre érkezik serkentés, hatásuk az erősítés miatt nagyobb lesz, mint a két bemenet egyszerű összege, emiatt ezt supra-lineáris összegződésnek nevezik. Olyan is lehetséges, hogy a dendrit egyik részére érkező bemenet csak akkor jut tovább, amikor a sejttesthez közelebbi dendritrészen beindulnak ezek az erősítő rendszerek. Így az egyik agyterületről származó jel csak akkor tud beterjedni, ha egy másik agyterületről megfelelő pillanatban szintén érkezik információ. Ezt az információ kapuzásának nevezik, hiszen az egyik bemenet nyitja meg a kaput a másik számára.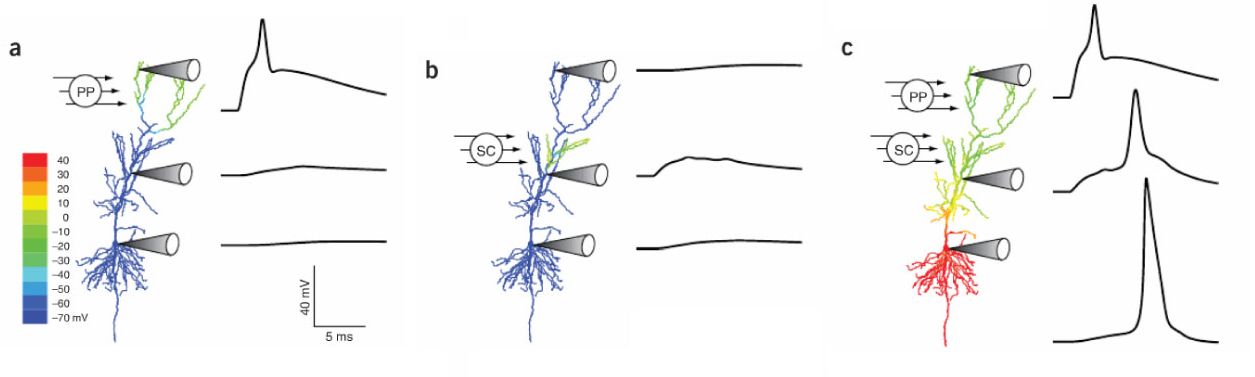
Amennyiben a sejt membránpotenciálját hiperpolarizáló K+ csatornákat kapcsolnak be a Ca2+ áramok vagy a membrán depolarizációja, akkor a beérkező serkentő bemenetek hatásának összege kisebb lesz, mint két bemenet összege a passzív esetben, ezért ezt sub-lineáris összegződésnek hívjuk.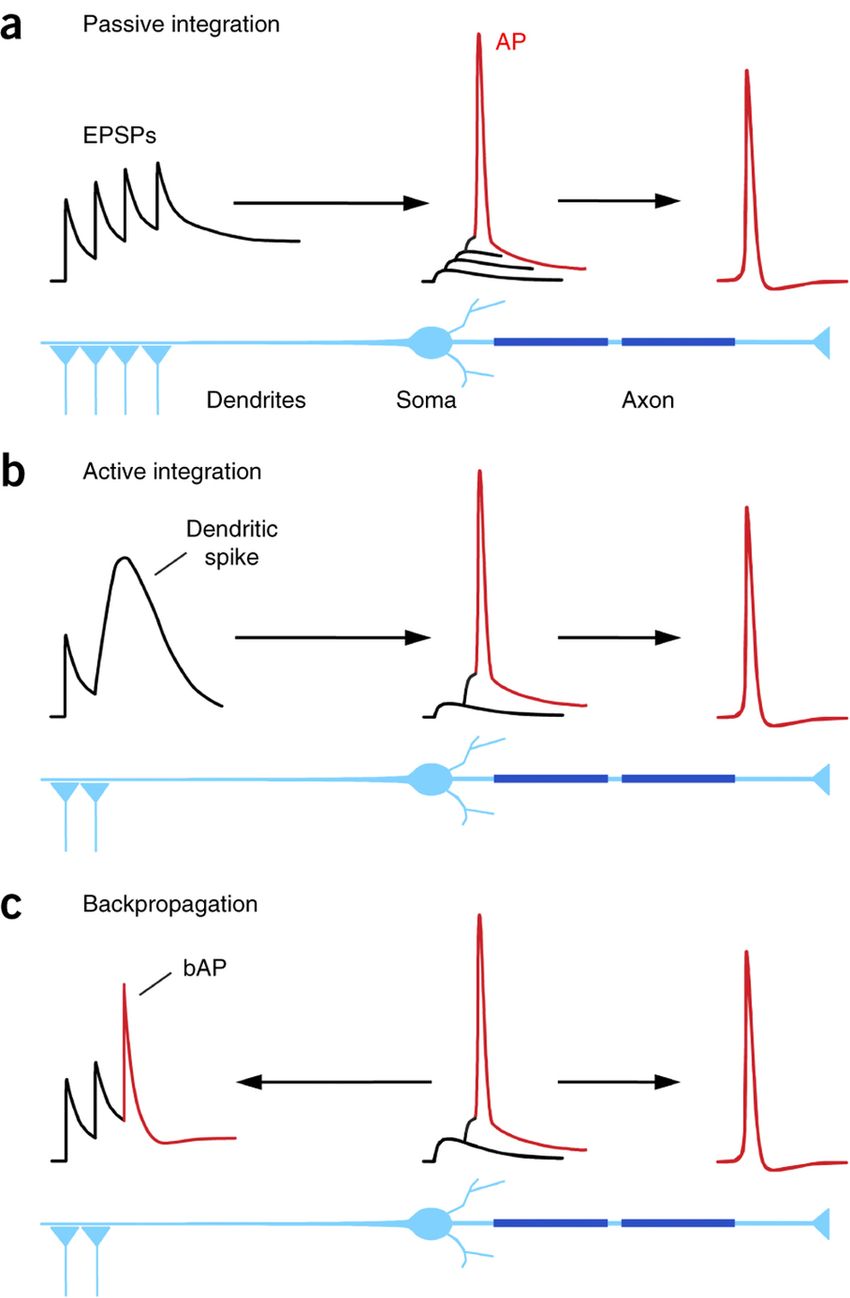 Hogy a sejtek jelfeldolgozó tulajdonságai még kifinomultabbak legyenek, bizonyos sejtek esetében a dendriteken is ki tudnak alakulni akciós potenciálok (AP), melyek terjedhetnek a dendritágak felől a sejttest felé vagy a sejttestről a dendritek felé. A két irányú terjedésnek eltérő szerepe van az összetett jelintegrációban.
Hogy a sejtek jelfeldolgozó tulajdonságai még kifinomultabbak legyenek, bizonyos sejtek esetében a dendriteken is ki tudnak alakulni akciós potenciálok (AP), melyek terjedhetnek a dendritágak felől a sejttest felé vagy a sejttestről a dendritek felé. A két irányú terjedésnek eltérő szerepe van az összetett jelintegrációban.
Amikor egy dendritágra elegendő mennyiségű serkentés érkezik ahhoz, hogy elérje a dendritikus akciós potenciál kialakulását, akkor a dendritről egy felerősített jel fut be a sejttestre. Ez lehet olyan erős is, hogy a sejtet is kisüsse. De az is előfordulhat, hogy több ilyen dendritikus akciós potenciálnak kell egyszerre beterjednie a sejtre ahhoz, hogy a sejt kisüljön. Mivel a dendritikus gátlás hatékonyan meg tudja vétózni egy dendriten az akciós potenciálok kialakulását, jelenléte vagy hiánya befolyásolja hogyan integrál egy sejt. A dendrit- és sejttest-gátlással, vagy a csatornák kapcsolgatásával, egyetlen idegsejt működhet úgy, hogy egyetlen dendritre érkező bemenet csoport kisütheti, de úgy is, hogy sok dendritágra sok bemenetnek kell egyszerre érkeznie ahhoz, hogy a sejt kisüljön. A sejt így állapot függően lehet logikai VAGY vagy ÉS kapu.
A dendriteken kialakuló AP-k gyakran Ca2+ áramoktól erednek. Ugye a Ca2+ szint fontos a szinaptikus súly módosulásánál. Ha egy bemenet Ca2+ akciós potenciált vált ki egy sejtben akkor az egész dendritben megemelkedik a Ca2+ szint és a dendritre érkező összes aktivált szinapszis esetében beindul a szinaptikus súlyok megerősödése.
Amennyiben a serkentő bemenetek dendritikus AP-t nem váltanak ki, de a sejttesten indítanak AP-t, akkor bizonyos sejttípusokban az AP a dendritekre is visszaterjedhet. Ennek itt megint abban van szerepe, hogy a szinapszisok számára a bekövetkező Ca2+ szint emelkedés jelezze, itt az idő tanulni, hacsak, egy jól elhelyezett gátlás ki nem kapcsolja ezt a visszaterjedő jelet. Az már csak extra csavar, hogy néha az egy vagy több dendritből beterjedő akciós potenciál visszapattan és a fenti módon visszaterjed az összes dendritbe.
Végezetül léteznek olyan, elsősorban K+ áramok is, melyek lassabban vagy gyorsabban a nyugalmi potenciál irányába igyekeznek húzni a sejtet, vagy akár ez alá húzzák azt. Ha ezek az áramok gyorsak, képesek a dendritekre érkező EPSPket megrövidíteni, ezzel pontosabbá tehetik a jelátvitelt. Lassabb K+ csatornák pedig a sejtek tüzelési küszöbének beállításában fontosak. Ez utóbbiak a rövidtávú memóriában is szerepet játszhatnak, azáltal, hogy a sejtre érkező bemenetek egy időre megváltoztatják a sejt tüzelési küszöbét, és ezek után egy ideig egyes sejtek könnyebben vagy nehezebben tudnak kisülni. Ez a mechanizmus a rövidtávú memóriában játszhat szerepet, egy információt hordozó mintázat sejtjei, aktiválódásuk után egy ideig könnyebben kapcsolnak be, a mintázat rövid ideig fennmarad.
Összefoglalva: a valós, bonyolult idegsejtekben számos olyan ioncsatorna van még jelen, amelyektől a sejtek membránjai aktívvá válnak, mely a jelek jobb vagy rosszabb hatékonyságú összegződését és terjedését eredményezi. A csatornák működésének hangolásával a sejt bemenetei között összetett kölcsönhatások alakulhatnak ki, illetve a szinaptikus tanulás hatékonysága is változhat.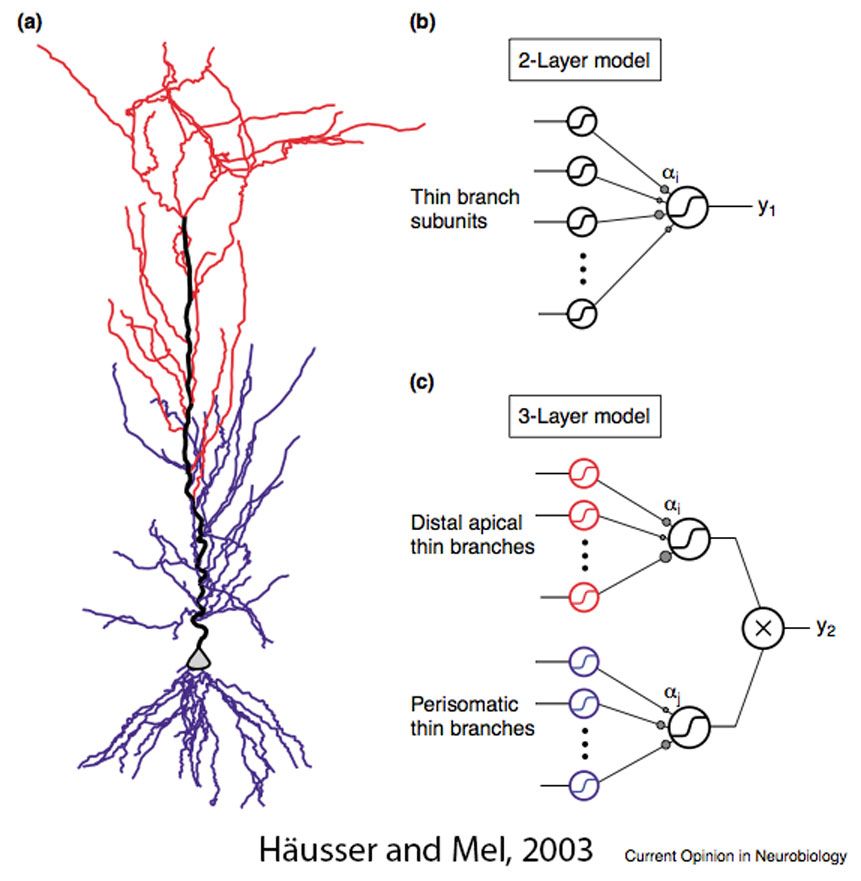 A legfontosabb hatás azonban az, hogy míg az egyszerű idesejt modellekben egy lépcsőben összegződnek a bemenetek és ha ez eléri a küszöböt akkor AP, kimenet keletkezik. Addig a valós, bonyolult idegsejtekben -elsősorban az agykérgi serkentő sejtekben, a piramis sejtekben- többlépcsős jelösszegződés figyelhető meg. Az első lépcsőben, ha megfelelő mennyiségű bemenet érkezik a távoli, vékonyabb, aktív dendritekre, akkor kialakulhat rajtuk dendritikus AP, mely a csúcsi dendrit segítségével a sejttestre terjed. Itt minden egyes dendritág tekinthető úgy, mint egy korábbi egyszerű, passzív idegsejt. A második lépcsőben pedig az egyes dendritágakról beérkező dendritikus AP-k összegződnek és ha elérik a sejttest tüzelési küszöbét, a sejt kisül. De az is lehet, hogy dendritcsoportok összegződnek csak a második lépcsőben és ezen csoportok eredménye újra összegződik egy harmadik lépésben. A dendritek és a sejttest gátlásával a lépcsők küszöbe állítható, és így elérhető az is, hogy nagyon kevés bemenet kisüssön egy sejtet, de az is, hogy nagyon sok bemenet kelljen ehhez. A visszaterjedő AP-k szabályozásával pedig az előzőektől valamelyest függetlenül kapcsolható a memória kialakulásában szerepet játszó dendritikus Ca2+ szint emelkedés.
A legfontosabb hatás azonban az, hogy míg az egyszerű idesejt modellekben egy lépcsőben összegződnek a bemenetek és ha ez eléri a küszöböt akkor AP, kimenet keletkezik. Addig a valós, bonyolult idegsejtekben -elsősorban az agykérgi serkentő sejtekben, a piramis sejtekben- többlépcsős jelösszegződés figyelhető meg. Az első lépcsőben, ha megfelelő mennyiségű bemenet érkezik a távoli, vékonyabb, aktív dendritekre, akkor kialakulhat rajtuk dendritikus AP, mely a csúcsi dendrit segítségével a sejttestre terjed. Itt minden egyes dendritág tekinthető úgy, mint egy korábbi egyszerű, passzív idegsejt. A második lépcsőben pedig az egyes dendritágakról beérkező dendritikus AP-k összegződnek és ha elérik a sejttest tüzelési küszöbét, a sejt kisül. De az is lehet, hogy dendritcsoportok összegződnek csak a második lépcsőben és ezen csoportok eredménye újra összegződik egy harmadik lépésben. A dendritek és a sejttest gátlásával a lépcsők küszöbe állítható, és így elérhető az is, hogy nagyon kevés bemenet kisüssön egy sejtet, de az is, hogy nagyon sok bemenet kelljen ehhez. A visszaterjedő AP-k szabályozásával pedig az előzőektől valamelyest függetlenül kapcsolható a memória kialakulásában szerepet játszó dendritikus Ca2+ szint emelkedés.
Azt, hogy minek a hatására kapcsolódnak a csatornák és a gátlósejtek eltérő információfeldolgozást okozó állapotba a következő bejegyzés szól.
Szerző: Gulyás Attila