



A gátlósejtek és a hálózatok aktivitásának modulálása
Aki átugrotta az előző bejegyzést, melyben alaposan cikornyáztuk az idegsejtek jelfeldolgozását annak legyen annyi elég, hogy a sejthártyában, az alapvető jeltovábbító és jelintegráló feladatokat ellátó alap Na+, K+, Cl- és Ca2+ csatornákon kívül számtalan, sejttípusokra jellemző, finomabb működést lehetővé tevő csatorna található, melyeket különböző állapotokba kapcsolva megváltoznak a sejtek jelfeldolgozó tulajdonságai és ezáltal a sejtekből alkotott hálózatok számos feldolgozási módba kapcsolhatnak.
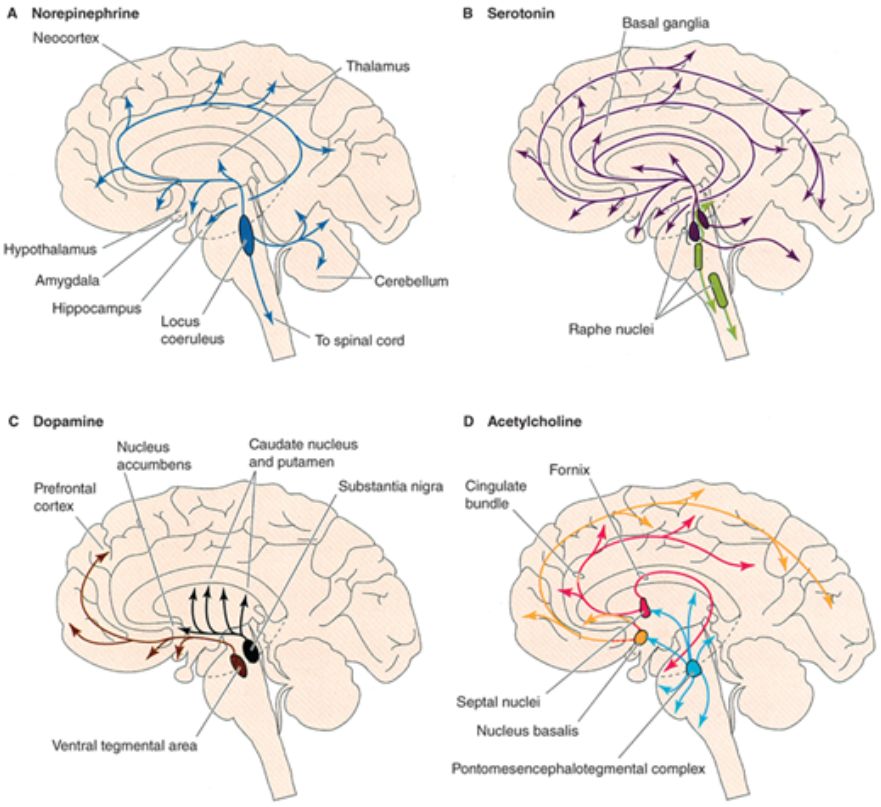
Az elmúlt évtized legfontosabb felismerése az volt, hogy az agyterületek megfelelő kölcsönhatásához az egyes területeket pontos időzítéssel kell a megfelelő működési állapotba kapcsolni. Az agy felépítésének bemutatásánál az agyat evolúciós története és felépítése alapján agykéregre és kéreg alatti területekre vágtuk. A kéreg alatti területekről felszálló moduláló (hangoló) pályák alapvető szerepet játszanak az agykérgi területek működésének hangolásában és összehangolásában, az állat viselkedési állapotának függvényében. A kéreg alatti területek a szervezet belső állapotáról, az állat motivációjáró, érzelmi állapotáról gyűjtenek információt, és ennek megfelelően kapcsolják az agykérgi területeket hatékonyan együttműködő feldolgozási állapotokba.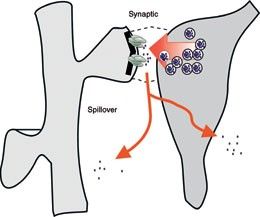 A felszálló szerotonint, hisztamint, dopamint, noradrenalint, acetil-kolint és GABAt felszabadító pályák változatos módokon hatnak az agykéreg serkentő és gátló sejtjeire. A pályakövető és mikroszkópos vizsgálatok megállapították, hogy ezekből a rendszerekből az átvivőanyagok felszabadulhatnak szinapszisok nélkül, nem célzottan, a sejtek közötti térbe, lassan hatva a sejtekre. De célzottan is felszabadulhatnak szinapszisok útján, eltérő típusú sejtekre hatva, jelentősen gyorsabban. A vizsgálatok szerint számos rendszer kiválogatja magának a gátlósejteket, azokon belül is különböző típusú gátlásokat létrehozó alcsoportjaikat és ezek működésének modulációja útján képes igen hatékonyan a serkentő sejtek jelintegrációját befolyásolni.
A felszálló szerotonint, hisztamint, dopamint, noradrenalint, acetil-kolint és GABAt felszabadító pályák változatos módokon hatnak az agykéreg serkentő és gátló sejtjeire. A pályakövető és mikroszkópos vizsgálatok megállapították, hogy ezekből a rendszerekből az átvivőanyagok felszabadulhatnak szinapszisok nélkül, nem célzottan, a sejtek közötti térbe, lassan hatva a sejtekre. De célzottan is felszabadulhatnak szinapszisok útján, eltérő típusú sejtekre hatva, jelentősen gyorsabban. A vizsgálatok szerint számos rendszer kiválogatja magának a gátlósejteket, azokon belül is különböző típusú gátlásokat létrehozó alcsoportjaikat és ezek működésének modulációja útján képes igen hatékonyan a serkentő sejtek jelintegrációját befolyásolni.
A serkentő sejtek jelfeldolgozását a moduláló rendszerek tehát két úton befolyásolják. Egyrészt szinaptikus vagy nem szinaptikus módon a serkentő sejtek felszínén elhelyezkedő ionotróp és metabotróp receptorokra hatva megváltoztatják bizonyos ioncsatornák működését és ezáltal a sejtek jelintegrációs tulajdonságait. Másrészt eltérő gátlósejt csoportokat megcélozva, hasonlóan módosítják azok jelintegrációját, melynek eredményeként megváltozik a serkentő sejtekre érkező gátlás. A két hatás befolyásolja a jelek összegzését, a szinaptikus súly megváltozását vagy a sejt küszöbének rövidebb-hosszabb idejű megváltozását.
 Intézetünk kutatói, köztük jómagam eredményei szerint például a mediális szeptumból eredő GABA tartalmú rostok válogatás nélkül az összes agykérgi gátlósejt csoport aktivitását csökkentik és ezért közvetve növelik a serkentő sejtek aktivitását. Ez a pálya fontos lehet abban, hogy amikor az állat felderíti a környezetét, a hippokampusz nevű agyterület felváltva tudjon tervező és információ gyűjtő állapotok között átkapcsolni. Míg egy másik, a szintén általunk vizsgált, szerotonint tartalmazó mediális raphe mag idegsejtjeiből induló pályarendszer az agykéregben csak a dendriteket beidegző gátlósejteken végződik, és feltételezhetőleg a sejtek tanulási rendszerét kapcsolja be vagy ki, a megfelelő magatartási állapotban, amikor tanulásra van szükség.
Intézetünk kutatói, köztük jómagam eredményei szerint például a mediális szeptumból eredő GABA tartalmú rostok válogatás nélkül az összes agykérgi gátlósejt csoport aktivitását csökkentik és ezért közvetve növelik a serkentő sejtek aktivitását. Ez a pálya fontos lehet abban, hogy amikor az állat felderíti a környezetét, a hippokampusz nevű agyterület felváltva tudjon tervező és információ gyűjtő állapotok között átkapcsolni. Míg egy másik, a szintén általunk vizsgált, szerotonint tartalmazó mediális raphe mag idegsejtjeiből induló pályarendszer az agykéregben csak a dendriteket beidegző gátlósejteken végződik, és feltételezhetőleg a sejtek tanulási rendszerét kapcsolja be vagy ki, a megfelelő magatartási állapotban, amikor tanulásra van szükség.
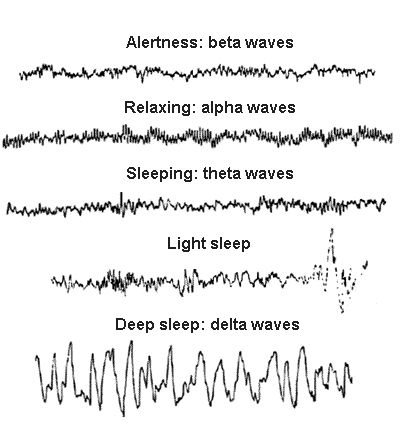 Ezek a gátlósejteken keresztül történő hangoló hatások felelősek azért, hogy az agykéreg egyes területei finoman összehangolt időzítéssel különböző EEG hullámokat mutató aktivitási módok között kapcsolgassanak. A figyelemhez társuló gamma, a pihenés alatt megjelenő alfa vagy az alvásra jellemző delta, theta aktivitások mindegyikében más moduláló ingerületátvivő közegben úszik az agy, amelyek hatására a serkentő és gátlósejtek jelintegrációs tulajdonságai az adott feladat elvégzésére alkalmas állapotba kerülnek.
Ezek a gátlósejteken keresztül történő hangoló hatások felelősek azért, hogy az agykéreg egyes területei finoman összehangolt időzítéssel különböző EEG hullámokat mutató aktivitási módok között kapcsolgassanak. A figyelemhez társuló gamma, a pihenés alatt megjelenő alfa vagy az alvásra jellemző delta, theta aktivitások mindegyikében más moduláló ingerületátvivő közegben úszik az agy, amelyek hatására a serkentő és gátlósejtek jelintegrációs tulajdonságai az adott feladat elvégzésére alkalmas állapotba kerülnek.
Szerző: Gulyás Attila